11
UN GRAND PAS POUR L’HUMANITÉ
Dans mon livre précédent, Darwin et les grandes énigmes de la vie, un de mes essais sur l’évolution humaine commence par ces mots :
« On a découvert un si grand nombre de fossiles humains au cours de ces dernières années que chaque année, quand arrive le moment de traiter ce sujet, j’ouvre mon vieux dossier et classe le contenu dans les archives. Et on recommence tout. »
Je me félicite chaudement de les avoir écrits car ils me permettent à présent de réfuter une thèse avancée dans ce même article.
J’y mentionnais en effet la découverte faite par Mary Leakey à Laetoli, à cinquante kilomètres au sud de la gorge d’Olduvai en Tanzanie, du plus vieux fossile d’hominidé connu – dents et mâchoires de 3,35 à 3,75 millions d’années. Mary Leakey déclarait alors (et pour autant que je sache, croit toujours) que ces restes devaient être classés dans notre genre Homo. J’en avais déduit que la lignée évolutive de l’homme que l’on faisait aller de l’australopithèque, doté d’un petit cerveau mais se tenant droit, à l’Homo au gros cerveau, devrait éventuellement être rectifiée, les australopithèques ne pouvant représenter qu’un rameau annexe de l’arbre généalogique de l’homme.
Dans les premiers jours de 1979, la presse annonça avec fracas la découverte d’une nouvelle espèce – plus ancienne et d’apparence plus primitive que tout autre hominidé fossile –, l’Australopithecus afarensis, ainsi dénommée par Don Johanson et Tim White. Était-il possible d’imaginer deux conceptions aussi radicalement opposées, celle de Mary Leakey pour qui les plus vieux hominidés appartenaient à notre genre Homo et celle de Johanson et de White qui avaient pris la décision de baptiser ainsi cette nouvelle espèce au vu de caractéristiques simiesques que ne possède aucun autre hominidé fossile ? Johanson et White ont-ils découvert des ossements nouveaux et fondamentalement différents ? Pas du tout. Le désaccord entre Leakey et Johanson-White porte sur les mêmes ossements. Nous sommes les témoins d’un débat sur une interprétation, non sur une nouvelle découverte.
Johanson a travaillé en Éthiopie dans la région de l’Afar de 1972 à 1977 et y a mis au jour une série exceptionnelle de restes d’hominidés. Ils ont été datés entre 2,9 et 3,3 millions d’années. Le plus remarquable de ces vestiges est le squelette d’une australopithèque baptisée Lucy. Il est complet à 40 %, ce qui est beaucoup plus que tout ce qu’on a pu posséder sur un seul individu de ces premiers temps de notre histoire. (La plupart des hominidés fossiles, bien qu’ils soient à la base de discussions sans fin et d’élucubrations laborieuses, ne sont que des fragments de mâchoires et des morceaux de crâne.)
Selon Johanson et White, les restes trouvés dans l’Afar et les fossiles de Laetoli découverts par Mary Leakey sont identiques quant à leur forme et appartiennent à la même espèce. Ils font également remarquer que les os et les dents de l’Afar et de Laetoli représentent tout ce que nous savons sur les hominidés qui ont plus de 2,5 millions d’années – tous les autres fossiles africains sont plus récents. Selon eux, les dents et les fragments de crâne ont en commun un ensemble de caractères que l’on ne retrouve pas dans les fossiles ultérieurs et qui rappellent les singes. C’est pour cela qu’ils ont classé les restes de l’Afar et de Laetoli dans une nouvelle espèce, A. afarensis.
Le débat ne fait que commencer, mais d’ores et déjà trois opinions s’affrontent. Certains anthropologues, s’appuyant sur des caractères différents, considèrent que les fossiles de l’Afar et de Laetoli appartiennent à notre genre Homo. D’autres pensent, avec Johanson et White, que ces fossiles sont plus proches de l’australopithèque d’Afrique orientale et australe – qui, lui, est moins âgé – que du genre Homo. Mais ils ne lui trouvent pas de différence suffisamment marquée pour justifier la création d’une nouvelle espèce et préfèrent inclure les fossiles de l’Afar et de Laetoli dans l’espèce A. africanus, nom donné originairement aux fossiles trouvés en Afrique du Sud dans les années 1920. D’autres encore s’accordent avec Johanson et White pour estimer que les fossiles de l’Afar et de Laetoli méritent une nouvelle dénomination. Moi-même n’étant qu’un profane dans le domaine de l’anatomie, mon avis n’est guère autorisé. Mais je dois dire que si une illustration vaut toutes les phrases de ce chapitre, le palais de l’hominidé de l’Afar m’apparaît bien comme celui d’un « singe ». (Je dois avouer aussi que l’appellation A. afarensis vient conforter plusieurs de mes préjugés favoris. Johanson et White soulignent que les fossiles de l’Afar et de Laetoli sont séparés par un espace de temps de 1 million d’années, mais qu’ils sont pratiquement identiques. Je pense que la plupart des espèces ne se modifient guère pendant la longue période de leur succès et que les transformations évolutives interviennent au cours de rapides événements durant lesquels les espèces s’écartent de leur souche ancestrale – voir les chapitres 17 et 18. En outre, puisque à mes yeux l’évolution humaine se présente davantage comme un buisson que comme une échelle, plus il y a d’espèces, mieux cela vaut. Johanson et White cependant acceptent une progressivité beaucoup plus grande que celle dont je serais partisan en ce qui concerne l’évolution humaine ultérieure.)
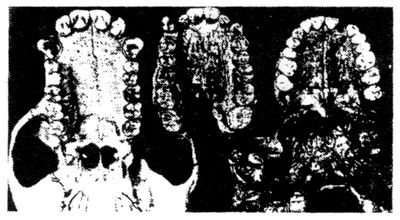
Le palais de l’Australopithecus
afarensis (au centre), comparé à celui
d’un chimpanzé actuel (à gauche) et à celui d’un humain (à
droite).
Avec l’aimable autorisation de Tim White et du Muséum d’histoire
naturelle de Cleveland.
Au cours de cette discussion portant sur le crâne, les dents et la classification taxonomique, une autre caractéristique des fossiles de l’Afar beaucoup plus intéressante n’a pas été prise en considération. Les os du bassin et de la jambe de Lucy montrent de toute évidence que l’A. afarensis marchait aussi droit que vous et moi. Ce fait a été longuement commenté dans la presse, mais d’une manière qui ne peut que prêter à confusion. Les journaux ont presque unanimement accrédité l’idée que, dans la pensée orthodoxe précédente, l’apparition d’un cerveau volumineux et de la station droite s’était faite de façon progressive et parallèle, avec peut-être une avance pour le cerveau. Cette transition allait de quadrupèdes à la cervelle minuscule à des êtres à la silhouette courbée et au cerveau à moitié développé jusqu’à l’Homo, parfaitement droit et doté d’un gros cerveau. Le New York Times a écrit en janvier 1979 : « On pensait que la bipédie avait été un processus graduel qui aurait inclus des hommes-singes, intermédiaires des êtres humains actuels, marchant courbés en traînant les pieds, créatures plus intelligentes que les singes, mais moins intelligentes que les êtres humains actuels. » Ce qui est absolument faux, au moins pour les cinquante dernières années de nos connaissances.
On sait, depuis les découvertes réalisées dans les années 1920, que les australopithèques avaient un cerveau peu développé et connaissaient la station verticale. (L’A. africanus avait un cerveau trois fois plus petit que le nôtre et marchait complètement droit. Même si l’on corrige sa capacité crânienne en fonction de sa taille réduite, son cerveau reste considérablement plus petit que le nôtre.) Cette « anomalie » que constitue la coexistence d’un cerveau réduit et de la station droite a fait couler beaucoup d’encre pendant des dizaines d’années et a occupé une place éminente dans tous les textes importants.
La dénomination d’A. afarensis n’établit donc pas la priorité historique de la station droite sur les cerveaux volumineux. Mais, conjointement avec deux autres idées, elle éclaire l’antériorité de la station droite d’un jour nouveau qui était resté curieusement absent dans les comptes rendus de la presse ou enfoui sous des masses d’informations erronées. L’A. afarensis est important car il nous apprend que la station verticale perfectionnée était déjà acquise il y a près de 4 millions d’années. La structure du bassin de Lucy montre qu’elle utilisait la bipédie dont les remarquables empreintes de pieds que l’on vient de découvrir à Laetoli apportent une preuve plus éclatante encore. Les australopithèques plus tardifs d’Afrique australe et orientale ne remontent guère à plus de 2,5 millions d’années. Nous avons ainsi ajouté près de 1,5 million d’années à l’histoire de la station droite.
Pour faire comprendre à quel point ce nouvel apport est important, il me faut abandonner le cours de mon exposé et me déplacer vers l’extrémité opposée de la biologie, c’est-à-dire aller des fossiles d’animaux complets aux molécules. Durant ces quinze dernières années, les spécialistes de l’évolution moléculaire ont accumulé une grande quantité d’informations sur les séquences des acides aminés d’enzymes et de protéines similaires chez de nombreux organismes variés. Ces données ont permis d’aboutir à des résultats surprenants. Si l’on prend deux espèces dont on connaît avec certitude la date à laquelle elles ont divergé d’un ancêtre commun, on remarque que les différences des acides aminés correspondent exactement au temps écoulé depuis la disjonction : plus la période de séparation est éloignée, plus la différence moléculaire est grande. Cette régularité a autorisé l’établissement d’une horloge moléculaire pour déterminer les dates de divergence chez des couples d’espèces pour lesquels les fossiles ont apporté la preuve d’une ascendance commune. Il est vrai que cette horloge ne bat pas avec la fiabilité d’une montre de haute précision – un de ses chauds partisans l’a même qualifiée de « tocante » – mais elle a rarement battu tout à fait la breloque.
Les darwiniens ont généralement été surpris par la régularité de l’horloge, car la sélection naturelle devrait normalement, selon les différentes lignées et les différentes périodes, travailler à des vitesses présentant des variations prononcées : très rapides dans les formes complexes s’adaptant à des milieux changeant fréquemment, très lentes au sein de populations stables et bien adaptées. Si la sélection naturelle est la cause principale de l’évolution au sein d’une population, on devrait alors s’attendre à une bonne corrélation entre le changement génétique et le temps, à moins que les vitesses de sélection ne demeurent assez constantes – comme cela ne devrait pas être le cas si l’on s’en réfère à l’argument exposé plus haut. Les darwiniens ont évité cette contradiction en affirmant que les irrégularités dans la vitesse de sélection s’aplanissaient sur de longues périodes. La sélection peut être très intense pendant quelques générations et pratiquement absente durant la période suivante sans que le changement global, enregistré sur de longues périodes, perde de sa régularité. Mais les darwiniens se sont vus dans l’obligation d’envisager une autre possibilité : la régularité de l’horloge moléculaire serait le reflet d’un processus évolutif que ne provoquerait pas la sélection naturelle, la fixation fortuite de mutations neutres. (Je dois reporter ce sujet brûlant à plus tard car il mérite un long développement.)
En tout cas, la mesure des différences d’acides aminés entre les humains et les grands singes d’Afrique (gorilles et chimpanzés) a apporté les plus surprenants des résultats. Pour les gènes qui ont été étudiés, nous sommes pratiquement identiques malgré notre divergence morphologique prononcée. La différence moyenne dans les séquences d’acides aminés entre l’homme et les singes africains est inférieure à 1 % (0,8 % pour être précis), ce qui, sur l’échelle moléculaire, correspond seulement à une période de 5 millions d’années depuis la divergence d’un ancêtre commun. En tenant compte de l’imprécision de cette « tocante » moléculaire, Allan Wilson et Vincent Sarich, les chercheurs de Berkeley qui ont découvert cette anomalie, sont prêts à accepter le chiffre de 6 millions d’années, mais guère plus. En bref, si l’horloge donne l’heure exacte, l’A. afarensis se retrouve très proche de la limite théorique de la branche des hominidés.
Jusqu’à une date récente, les anthropologues tendaient plutôt à refuser cette horloge car, selon eux, les hominidés constituaient une véritable exception à une règle admise. Ils fondaient leurs réticences sur l’existence d’un animal appelé Ramapithecus, fossile africain et asiatique dont on ne connaît guère que quelques fragments de mâchoires et qui vivait voici environ 14 millions d’années. Pour de nombreux anthropologues, le ramapithèque pouvait se placer de notre côté de la disjonction singe-homme, ce qui signifiait en d’autres termes que la divergence entre les hominidés et les singes s’était produite il y a plus de 14 millions d’années. Mais cette thèse qui s’appuie sur une série d’arguments techniques – les dents et leurs proportions – a récemment perdu du terrain. Certains des partisans les plus ardents du ramapithèque comme hominidé sont à présent prêts à réexaminer leur position et à en faire un singe ou une créature proche de la descendance commune aux singes et aux humains, mais cependant antérieure à la divergence. L’horloge moléculaire s’est trouvée vérifiée trop souvent pour qu’on la rejette pour quelques arguments sujets à révision fondés sur des fragments de mâchoires. (Je sens que je vais bientôt perdre un pari de dix dollars que j’ai fait avec Allan Wilson il y a quelques années. Généreusement il m’avait accordé 7 millions d’années comme l’âge maximal du plus vieil ancêtre commun aux singes et aux hommes alors que je penchais pour un chiffre plus élevé. Je n’ai pas encore payé, mais je ne m’attends pas vraiment à empocher de l’argent cette fois-ci{7}.)
Nous pouvons à présent rassembler les trois éléments qui vont nous permettre d’envisager une réorientation importante des thèses sur l’évolution humaine : l’âge et la station verticale de l’A. afarensis, la divergence singe-homme sur l’horloge moléculaire et la perte du titre d’hominidé par le ramapithèque.
Nous n’avons jamais pu nous débarrasser de cette vision de l’évolution humaine centrée sur le cerveau, bien que celle-ci n’ait jamais représenté autre chose qu’un puissant préjugé culturel plaqué que la nature. Les premiers évolutionnistes pensaient que le développement de la taille du cerveau devait avoir précédé toutes les modifications importantes de notre corps (voir les thèses de Grafton Elliot Smith dans le chapitre 10. Smith fondait sa conviction en faveur de l’homme de Piltdown sur une croyance presque fanatique en la priorité de l’encéphale). Mais l’A. africanus, avec sa station verticale et son cerveau réduit, mit fin à cette conception dans les années 1920, comme l’avaient prévu avec beaucoup de clairvoyance certains évolutionnistes et philosophes, d’Ernst Haeckel à Friedrich Engels. Néanmoins, cette « priorité cérébrale », comme je me plais à l’appeler, n’a pas lâché prise, mais a adopté une forme modifiée. Certains évolutionnistes, tout en admettant la primauté historique de la station verticale, ont supposé qu’elle était intervenue sans hâte et que la réelle discontinuité – le saut qui nous rendit réellement humains – se produisit beaucoup plus tardivement, lorsque, dans une explosion évolutive sans précédent, notre cerveau tripla de volume en à peu près 1 million d’années.
Voyons ce qu’écrivait un spécialiste éminent il y a dix ans : « Le grand saut dans la céphalisation du genre Homo a eu lieu il y a 2 millions d’années, après quelque 10 millions d’années d’évolution préparatoire à travers la bipédie, la main préhensile, etc. » Dans son dernier livre, Janus, Arthur Koestler a porté cette thèse à des sommets de divagation erronée rarement atteints. Notre cerveau a grossi si vite, d’après lui, que le cortex extérieur, siège de l’astuce et de la rationalité, a perdu le contrôle des centres animaux émotifs situés au plus profond de notre cerveau. Cette bestialité primitive resurgit dans la guerre, l’assassinat et les autres formes de violence destructrice.
Je pense qu’il nous faut revoir de fond en comble la place accordée jusqu’à présent à la station droite et au développement de la taille du cerveau dans l’évolution humaine. On a considéré que la station droite avait été une tendance progressive ayant aisément atteint son terme et que l’augmentation du volume du cerveau avait pris la forme d’une rupture étonnamment rapide, c’est-à-dire aurait été quelque chose d’exceptionnel tout à la fois dans son mode d’évolution et dans l’ampleur de son effet. Je souhaite présenter la thèse diamétralement opposée. C’est la station verticale qui a été la surprise, l’événement difficile, la reconstruction rapide et fondamentale de notre anatomie. L’accroissement de la taille du cerveau n’a été, en termes d’anatomie, qu’un épiphénomène secondaire, une transformation facile s’inscrivant dans le schéma général de l’évolution humaine.
Il y a tout au plus 6 millions d’années, si l’horloge moléculaire donne l’heure exacte (et Wilson et Sarich penchent plutôt pour cinq), nous partagions avec les gorilles et les chimpanzés notre dernier ancêtre commun. On peut penser que cette créature a d’abord marché à quatre pattes, mais qu’elle devait se déplacer occasionnellement sur ses deux jambes, comme le font de nombreux singes aujourd’hui. Un peu plus de 1 million d’années plus tard, nos ancêtres étaient aussi bipèdes que vous et moi. C’est cela et non le grossissement ultérieur de notre cerveau qui a constitué la grande étape de l’évolution humaine.
La bipédie n’est en rien une performance aisée. Elle requiert une restructuration fondamentale de notre anatomie, en particulier du pied et de la hanche. En outre, cette reconstitution anatomique s’écarte du schéma général de l’évolution humaine. Comme je l’explique dans le chapitre 9 par l’entremise de Mickey, les humains sont néoténiques, c’est-à-dire que nous nous sommes développés en conservant les caractéristiques juvéniles de nos ancêtres. Notre gros cerveau, notre petite mâchoire et de nombreux autres caractères, qui vont de la répartition de la pilosité à la disposition du vagin tourné vers le ventre, sont des conséquences de notre jeunesse éternelle. Mais la station droite est un phénomène différent. On n’a pas pu y parvenir en se contentant de conserver un caractère déjà présent dans les phases juvéniles. Car les jambes de bébé sont relativement petites et faibles, alors que la station droite exige que les jambes deviennent plus grosses et plus solides.
Le temps que nous devenions droits comme l’était l’A. afarensis, la partie était déjà jouée, les modifications essentielles de notre architecture avaient été accomplies, les éléments nécessaires pour les changements à venir étaient en place. Le développement ultérieur de notre cerveau fut anatomiquement facile. Il s’explique par le programme même de notre propre croissance : les vitesses rapides de la croissance fœtale y ont été reportées à une période ultérieure et les proportions qui caractérisent le crâne d’un jeune primate ont été conservées à l’âge adulte. Le cerveau s’est développé de concert avec d’autres traits néoténiques, tous inscrits dans un schéma évolutif général.
Mais avant de clore ce chapitre, il me faut revenir en arrière pour éviter une erreur de raisonnement, à savoir la fausse équation entre l’ampleur de l’effet et l’intensité de la cause. En tant que simple problème de reconstruction architecturale, la station verticale est fondamentale et d’une grande portée, la taille du cerveau est superficielle et secondaire. Mais l’effet de notre cerveau volumineux a surpassé de beaucoup la relative facilité de sa construction. La chose la plus surprenante de tout ceci est peut-être cette propriété générale des systèmes complexes, de notre cerveau en premier lieu, consistant à transformer de simples changements structurels quantitatifs en qualités fonctionnelles merveilleusement différentes.
Il est maintenant deux heures du matin et j’ai terminé mon chapitre. Je pense que je vais aller prendre une bière dans le réfrigérateur, puis j’irai me coucher. La créature que je suis, produit de la culture, s’étonnera toujours beaucoup plus du rêve qu’elle va avoir dans une heure en position horizontale que des quelques pas qu’elle va faire perpendiculairement au plancher.